К содержанию учебника «Антропология» | К следующей главе
В настоящее время наука располагает значительным числом палеоантропологических, археологических и геологических данных, позволяющих осветить ход антропогенеза (в общих чертах). Анализ указанной информации дает основание выделить четыре условных стадии (отрезка) антропогенеза, характеризующиеся определенным типом ископаемого человека, уровнем развития материальной культуры и общественных институтов:
1) австралопитеки (предшественники человека);
2) питекантропы (древнейшие люди, архантропы);
3) неандертальцы (древние люди, палеоантропы);
4) человек современного типа, ископаемый и современный (неоантропы).
В соответствии с зоологической систематикой классификация гоминид выглядит следующим образом:
Семейство — Hominidae
Подсемейство Australopithecinae — Австралопитеки
Род Australopithecus — Австралопитек
A. afarensis — А. афарский A. robustus — А. мощный A. boisei — А бойсов и др.
Подсемейство Homininae — Люди
Род Homo — Человек
Н. erectus — Человек выпрямленный
Н. sapiens neanderthalensis — Человек неандертальский разумный
Н. sapiens sapiens — Человек разумный разумный.
Автралопитеки (предшественники человека)
Палеонтологические и современные биологические (в большей степени) данные подтвердили теорию Дарвина о происхождении человека и современных антропологов от общей исходной формы.
Установление конкретного предка гоминоидов остается проблемой для современной науки. Существование его связано с большой, процветавшей в миоцене — плиоцене (миоцен простирается в пределах 22-27 млн лет, плиоцен — в пределах 5-10 млн лет) группой африканских дриопитеков, ведущей начало от олигоценового египтопитека (30 млн лет). В 50-60-х гг. один из дриопитеков — проконсул — выдвигался на роль «модели» общего предка гоминид и понгид. Миоценовые дриопитеки были полуназемными-полудревесными человекообразными обезьянами, обитавшими в условиях влажных тропических, горных или обычных широколиственных лесов, а также районов лесостепи. Находки миоценовых и нижнеплиоценовых дриопитеков известны также в Греции, Венгрии и Грузии.
От общей исходной формы разошлись две ветви эволюции: первая, понгидная, привела спустя много миллионов лет к современным человекообразным обезьянам, вторая, гоминидная, — к появлению, в конечном счете, человека современного анатомического типа. Эти две ветви в течение многих миллионов лет развивались независимо друг от друга в разных адаптивных направлениях. В соответствии с природными и ландшафтными условиями, в каждой из них сформировались специфические черты биологической организации, соответствующие образу жизни.
Ветвь высших обезьян эволюировала в направлении приспособления к древесному образу жизни, к локомоции брахиаторного типа со всеми вытекающими отсюда анатомическими особенностями: удлинением передних и укорочением задних конечностей, уменьшением большого пальца кисти, удлинением и сужением костей таза, развитием гребней на черепе, резким преобладанием лицевого отдела черепа над мозговым и т. д.
Человеческая ветвь эволюции, напротив, развивалась в направлении адаптации к наземному образу жизни, прямохождению, освобождению передних конечностей от функции опоры и локомоции, использованию их для употребления природных предметов в качестве орудий, а в дальнейшем — к изготовлению искусственных орудий, что и было решающим в выделении человека из природного мира. Выполнение этих задач требовало удлинения нижних и укорочения верхних конечностей, при этом стопа утрачивала хватательные функции и превращалась в орган опоры выпрямленного тела, бурно развивался головной мозг, главный, координирующий мозговой орган, и соответственно отдел черепа становится преобладающим; отмечается исчезновение гребней, надглазничного валика, образование подбородочного выступа на нижней челюсти и др.
Следующий важный вопрос эволюционной антропологии: когда возникла независимая ветвь эволюции человека и кто был первым ее представителем? Усреднение оценок, полученных палеонтологами и генетиками, дает нам период 8-6 млн лет. Генетики рассчитывают время разделения двух ветвей эволюции на основании генетических различий современных гоминоидов и предполагаемого времени его возникновения.
В качестве возможных предков гоминид, кроме рамапитека (последнего часто считают звеном в эволюции орангутангов), называются европейские высшие приматы: рудапитек и уранопитек, африканский кениапитек (потомок более древних проконсулов из «круга дриопитеков»), люфенгопитек (китайский рамапитек).
Австралопитеки представляют собою одну из первых стадий эволюции человека. Они могут рассматриваться наиболее осторожными исследователями как предшественники всех ископаемых и современных людей. Австралопитеки — наиболее интересный объект в современной палеонтологии человека — стали известны науке с 30-х годов нашего столетия. Первая находка австралопитека была сделана на юге Африканского материка. Она представляла собою остатки черепа и естественный отлив его мозговой части, принадлежащего детской особи.
Анализ «детеныша из Таунга» показал, что целый ряд структурных признаков отличается от типа антропоидов и одновременно походит на современного человека. Находка вызвала много споров: одни причисляли ее к ископаемым антропоидам, другие — к ископаемым гоминидам. Последующие находки южноафриканских австралопитеков продемонстрировали наличие двух морфологических типов — грациального и массивного австралопитеков. Первоначально они относились к двум самостоятельным родам. В настоящее время известно несколько сотен африканских австралопитеков. Южно- и восточноафриканские массивные и грациальные варианты австралопитеков отнесены к разным видам. Южноафриканские виды обитали в промежутке 3-1 млн лет, а восточноафриканские — 4 и более — 1 млн лет.
Современные антропологи не сомневаются в том, что австралопитеки — это промежуточный тип между человекообразными обезьянами и человеком. Основное отличие от первых — двуногое передвижение, отразившееся на строении скелета туловища и некоторых признаках черепа (срединное положение затылочного отверстия). Большая ширина тазовых костей, связанная с прикреплением ягодичных и части спинных мышц, выпрямляющих туловище, доказывает вертикальное положение туловища. К скелету таза прикрепляется также часть брюшных мышц, поддерживающих внутренние органы при ходьбе с выпрямленным корпусом.
Ландшафтное окружение австралопитеков — степь и лесостепь — требовало развития способности к передвижению на двух ногах. Иногда антропоиды демонстрируют такую способность. Для австралопитеков бипедия была постоянной особенностью. Экспериментально доказано, что двуногая походка энергетически более выгодна, чем остальные типы локомоции приматов.
Признаки человека современного типа выявлены на нижних челюстях. Относительно небольшие клыки и резцы не выступают над общим уровнем зубов. Довольно крупные коренные зубы имеют «человеческий» узор бугорков на жевательной поверхности, именуемый как «узор дриопитека». Строение зубов и сустава нижней челюсти свидетельствуют о преобладании в акте жевания боковых движений, что не свойственно антропоидам. Челюсти австралопитека массивнее, чем у современного человека. Близки к человеческому типу вертикальный профиль лицевого отдела и его относительно небольшая общая величина. Надбровье выступает вперед; мозговая полость небольшая; затылочный отдел имеет тенденцию к закруглению.
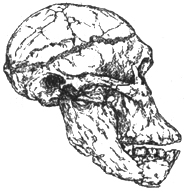
Рис. I. 5. Череп австралопитека африканского (2,5 млн лет)
Объем мозговой полости австралопитеков небольшой: грациальные австралопитеки — в среднем 450 см3, массивные австралопитеки — 517 см3, антропоиды — 480 см3, т. е. почти втрое меньше, чем у современного человека: 1450 см3. Таким образом, прогресса в развитии мозга по признаку абсолютной величины мозга в типе австралопитека практически не видно. Относительная величина мозга австралопитеков, в ряде случаев, была больше, чем у антропоидов.
Среди южноафриканских форм отчетливо выделяются «австралопитек африканский» и «австралопитек мощный». Последний можно охарактеризовать следующим образом: коренастое существо с длиной тела 150-155 см и весом около 70 кг. Череп более массивный, чем у австралопитека африканского, нижняя челюсть сильнее. Выраженный костный гребень на темени служили для прикрепления сильной жевательной мускулатуры. Зубы крупные (по абсолютным размерам), особенно коренные, при этом резцы непропорционально малы, так что хорошо видна диспропорция зубов. Такие морфологические особенности имел австралопитек-вегетарианец, тяготеющий в своем обитании к линии леса.
Австралопитек африканский имел меньшие размеры (грациальная форма): длина тела — до 120 см, а вес — до 40 кг (рис. I. 5). Судя по костям туловища, положение тела при ходьбе было более выпрямленным.
Структура зубов соответствовала адаптации к всеядности с большим удельным весом мясной пищи. Австралопитеки занимались собирательством и охотой, возможно, пользуясь при этом охотничьими трофеями других хищников. Охотясь на павианов, австралопитеки использовали в качестве метательного оружия камни. Р. Дарт создал оригинальную концепцию предкультуры австралопитеков — «остеодонтокератической культуры», т. е. постоянного использования частей скелета животных как орудий. Было высказано предположение об усложнении умственной деятельности австралопитеков: об этом свидетельствовали высокий уровень их орудийной деятельности и развитая стадность. Предпосылками этих достижений были прямохождение и развивающаяся рука.
Интересны находки австралопитеков и близких к ним форм, сделанные в Восточной Африке, в частности, в Олдувайском ущелье (Танзания). Антрополог Л. Лики проводил здесь исследования на протяжении 40 лет. Он выделил пять стратиграфических слоев, которые позволили установить временную динамику древнейших гоминид и их культуры в раннем плейстоцене.
Первоначально в Олдувайском ущелье был открыт череп массивного австралопитека, названного «Зинджантроп бойсов» («Щелкунчик»), позднее переименованного в «Австралопитека бойсова». Эта находка приурочена к верхней половине слоя I (возраст 2,3-1,4 млн лет). Примечательны найденные здесь же архаичные каменные орудия в виде отщепов со следами ретуши. Исследователи были смущены сочетанием каменной культуры и примитивного морфологического типа австралопитека. Позднее в слое I ниже зинджантропа были найдены кости черепа и руки более прогрессивного по типу человеческого существа. Именно ему, так называемому Homo habilis (Человеку умелому) и принадлежали олдувайские древнейшие орудия.
Что касается зинджантропа (A. boisei), то в эволюции австралопитеков он продолжает линию адаптации массивных форм к преимущественному питанию растительной пищей. Данный австралопитек крупнее «австралопитека мощного» и отличается менее совершенной способностью к двуногому хождению (рис. I. 6).
Большое значение имеет доказанный ископаемыми материалами Олдувайского ущелья факт сосуществования двух типов ранних гоминид — Австралопитека бойсова и Homo habilis, тем более, что они весьма заметно отличаются по морфологии и способам адаптации.
Остатки хабилиса в Олдувайском ущелье не единичны: они всегда соседствуют с галечной (олдувайской) культурой, древнейшей культурой палеолита. Некоторые антропологи оспаривают родовое название
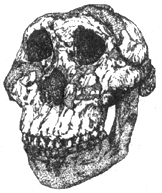
Рис. I. 6. Череп сверхмассивного австралопитека («бойсова») (1,9 млн лет)
хабилиса — «Ноmо», предпочитая называть его «австралопитеком умелым». Для большинства специалистов хабилис — древнейший представитель рода Homo. Он не только использовал для своих нужд подходящие предметы окружающей природы, но и видоизменял их. Древность Homo habilis — 1,9 — 1,6 млн лет. Находки данного гоминида известны в Южной и Восточной Африке.
Homo habilis имел длину тела до 120 см, при весе до 40-50 кг. Структура челюсти выдает его способность к всеядности (особенность человека). От зинджантропа хабилиса отличается большим объемом мозговой полости (объем — 660 см3), а также выпуклость свода черепа, особенно в затылочной области. Нижняя челюсть хабилиса грациальнее, чем у других австралопитеков, зубы мельче. В связи с довольно совершенным двуногим хождением большой палец стопы мог двигаться, как у человека, только в вертикальном направлении, а стопа обладала сводчатостью. Тело хабилиса было практически выпрямленным. Таким образом, бипедия как одно из основных достижений антропогенеза оформилась очень рано. Рука изменялась медленнее. Отсутствует совершенное противопоставление большого пальца остальным, размеры его, судя по костным элементам, малы. Фаланги пальцев изогнуты, что не характерно для человека современного типа, но концевые фаланги плоские.
В слоях Олувайского ущелья (возраст от 1,2-1,3 млн лет) найдены костные остатки форм, которые можно трактовать как переходные от типа прогрессивных австралопитеков к типу питекантропов. В данном местонахождении открыт и питекантроп.
Трудно поддаются интерпретации и классификации формы, сходные с австралопитеками Африки, но найденные вне данного материка. Так, на острове Ява открыт фрагмент нижней челюсти высшего примата, общие размеры которого значительно превышали размеры современного человека и наиболее крупных обезьян. Он получил название «Мегантроп палеояванский». В настоящее время его часто относят к группе австралопитеков.
Всем указанным австралопитекам и ранним представителям рода Homo предшествовали во времени грациальные «афарские австралопитеки» (A. afarensis), костные остатки которых открыты в Эфиопии и Танзании. Древность представителей данного вида — 3,9-3,0 млн лет. Счастливая находка весьма полного скелета субъекта, получившего имя «Люси», позволяет представить афарских австралопитеков следующим образом. Размеры тела весьма невелики: длина тела — 105-107 см, вес несколько превышал 29 кг. В строении черепа, челюстей и зубов отмечены очень примитивные признаки. Скелет адаптирован к двуногой походке, хотя и отличной от человеческой. Изучение отпечатков стоп в вулканическом пепле (древность — не менее 3,6 млн лет) приводит к выводу, что афарские австралопитеки не полностью разгибали ногу в тазобедренном суставе, а при ходьбе перекрещивали стопы, ставя их одну впереди другой. Стопа сочетает прогрессивные особенности (крупный и приведенный первый палец, выраженный свод, сформированная пятка) и обезьяноподобные черты (предплясна не неподвижна). Пропорции верхних
и нижних конечностей соответствуют прямохождению, но при этом есть явные признаки адаптации к древесному способу передвижения. В кисти также прогрессивные признаки сочетаются с архаичными (относительная укороченность пальцев), связанными со способностью к древесному передвижению. Признаков «силового захвата», характерных для гоминид, не наблюдается. Как примитивные черты черепа следует отметить сильное выступание лицевого отдела и развитый затылочный рельеф. Архаично даже на фоне других австралопитеков выглядят выступающие клыки и диастемы между зубами верхних и нижних челюстей. Очень крупны и массивны коренные зубы. Абсолютная величина мозга «афарского австралопитека» не отличима от величины антропоморфных обезьян, но его относительная величина несколько больше. Отдельные особи афарцев имеют четкую «шимпанзоидную» морфологию, доказывающую не столь отдаленное разделение эволюционных ветвей гоминид и понгид.
Некоторые неврологи считают, что у весьма древних представителей австралопитековых уже можно зафиксировать структурную перестройку теменных, затылочных и височных областей мозга; в то же время, помимо других, внешняя морфология мозга не отличима от обезьяньей. Перестройка мозга могла начинаться на клеточном уровне.
Самые современные палеоантропологические открытия позволяют предварительно выделить вид австралопитеков, во времени предшествовавших «афарцам». Это — восточноафриканский австралопитек A. ramidus (Эфиопия) (представлен нижней челюстью) и A. anamensis (Кения); (представлен фрагментами жевательного аппарата). Древность обеих находок около 4 млн лет. Имеются и более древние находки австралопитеков, не имеющие видового определения. Они заполняют временной хиатус между древнейшими австралопитеками и гоминоидным предком.
Большой интерес представляют находки ранних представителей рода Homo, сделанные на восточном берегу оз. Туркана (Кения). К прогрессивным признакам Homo habilis «1470» относятся объем мозга около 770 см3 и сглаженный рельеф черепа; древность — около 1,9 млн лет.
Какое место занимала орудийная деятельность в эволюционных достижениях австралопитеков? У антропологов нет единого мнения относительно нерасторжимости связи орудийной деятельности и двуногого хождения. Несмотря на находки очень древних культур каменных орудий, существует значительный разрыв во времени между появлением двуногости и возникновением труда. Предполагается, что причиной выделения первых гоминид из животного мира мог стать перенос оборонительной функции зубного аппарата на искусственные орудия защиты, а употребление орудий стало эффективной адаптацией в поведении первых людей, заселивших саванну. Памятники олдувайской культуры не прояснили вопроса о связи австралопитеков с олдувайскими орудиями. Так, известен факт находки костей прогрессивного «хабилиса» и массивного австралопитека в одном горизонте с олдувайскими орудиями.
Древнейшие орудия найдены в более древних горизонтах, чем фрагменты первых бесспорных представителей рода Homo. Так, палеолитические культуры в Кении и Эфиопии имеют возраст в 2,5-2,6 млн лет. Анализ новых материалов показывает, что австралопитеки были способны лишь к использованию орудий, но выделывать их умели лишь представители рода Homo.
Олдувайская (галечная) эпоха — самая ранняя в палеолите (древнем каменном веке). Наиболее характерные орудия — массивные архаичные поделки из галек и обломков камня, а также камни — заготовки (нуклеусы), орудия на отщепах. Типичное орудие олдувая — чоппер. Он представлял собой гальку со стесанным концом, необработанная часть которой служила для удержания орудия в руке (рис. I. 7). Лезвие могло быть обработано с двух сторон; найдены также орудия с несколькими гранями и просто ударные камни. Олдувайские орудия отличаются по форме и величине, но имеют одинаковый тип лезвия. Это объясняется целенаправленностью действий по выработке орудий. Археологи отмечают, что уже с начала палеолита существовал набор орудий разного назначения. Находки разбитых костей позволяют думать, что австралопитеки были охотниками. Олдувайские орудия доживают до поздних времен, особенно в Южной и Юго-Восточной Азии. Длительное существование Олдувая (1,5 млн лет) почти не сопровождалось техническим прогрессом. Австралопитеки могли устраивать простейшие укрытия типа ветровых заслонов.
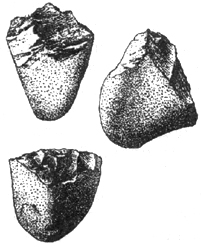
Рис. I. 7. Олдувайская культура нижнего палеолита. Питекантропы
(древнейшие люди, архантропы)
Питекантропы — это вторая по счету стадиальная группа гоминид после австралопитековых. В этом аспекте в специальной литературе они часто обозначаются (все варианты группы) как «архантропы», т. е. «древнейшие люди»; сюда можно также добавить определение «истинные люди», так как принадлежность питекантропов к семейству гоминид никем из антропологов не оспаривается. Ранее некоторые исследователи объединяли питекантропов с неандертальцами в одну эволюционную стадию.
Находки питекантропов известны в трех частях света — Африке, Азии и Европе. Их предками были представители Homo habilis (поздние восточноафриканские представители данного вида часто именуются Homo rudolfensis). Время существования питекантропов (включая самых ранних, Homo ergaster) можно представить в интервале 1,8 млн лет — менее 200 тыс. лет. Наиболее древние представители стадии открыты в Африке (1,6 млн лет — 1,8 млн лет); с рубежа 1 млн лет они распространены в Азии, а со времени 0,5 млн лет питекантропы (именуются часто «пренеандертальцы», или представители Homo heidelbergensis) обитали в Европе. Почти всесветное распространение питекантропов можно объяснить их достаточно высоким уровнем биологического и социального развития. Эволюция различных групп питекантропов происходила с различной скоростью, но имела одно направление — к сапиентному типу.
Впервые костные фрагменты питекантропа обнаружил голландский врач Е. Дюбуа на о. Ява в 1891 г. Примечательно, что автор находки разделял концепцию «промежуточного звена» в родословной человека, принадлежавшей дарвинисту Э. Геккелю. Вблизи села Триниль были найдены (последовательно) верхний коренной зуб, черепная крышка и бедренная кость. Впечатляет архаичный характер черепной крышки: покатый лоб и мощный надглазничный валик и вполне современный тип бедренной кости. Слои, содержащие тринильскую фауну, датируются временем 700 тыс. лет (в настоящее время — 500 тыс. лет). В 1894 г. Г. Дюбуа впервые дал научное описание «Питпекантропуса эректуса» («обезьяночеловека прямоходящего»). Столь феноменальную находку некоторые европейские ученые встретили в недоверием, да и сам Дюбуа часто не верил в ее значение для науки.
С интервалом в 40 лет были сделаны другие находки питекантропов на о. Ява и в других местонахождениях. В слоях Пунгата с фауной джетис у селения Моджокерто открыт детский череп питекантропа. Возраст находки близок к 1 млн лет. Находки костей черепа и скелета сделаны в местонахождении Сангиран (древность около 800 тыс. лет) в течение 1936-1941 гг. Следующая серия находок у Сангирана относится к периоду 1952-1973 гг. Наиболее интересна находка черепа питекантропа с сохранившимся лицевым отделом черепа, сделанная в 1963 г. Остатков палеолитической культуры на о. Ява не обнаружено.
Ископаемый человек сходного с питекантропом типа обнаружен в среднеплейстоценовых отложениях Китая. Зубы синантропа (китайского питекантропа) были обнаружены в известняковой пещере Чжоукоу-дянь в 1918 г. Сборы случайных находок сменились раскопками, и в 1937 г. в данном местонахождении были открыты остатки более 40 индивидуумов синантропа (рис. 1.8). Описание данного варианта питекантропов впервые сделал канадский специалист Влеком. Абсолютная датировка синантропа оценивается в 400-500 тыс. лет. Костные остатки синантропа сопровождаются многочисленными культурными
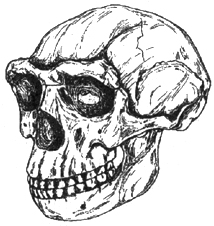
Рис. I. 8. Череп китайского питекантропа (0,4 млн лет)
остатками (каменные орудия, раздробленные и обожженные кости животных). Наибольший интерес представляет многометровая толща золы, найденная в охотничьем лагере синантропов. Применение огня для обработки пищи делало ее более усвояемой, а длительное поддержание костра свидетельствует о достаточно высоком уровне развития общественных связей у синантропов.
Множественные находки позволяют уверенно говорить о реальности таксона питекантропов. Приведем основные черты его морфотипа. Современный тип бедренных костей и положение большого затылочного отверстия, сходное с тем, что мы видим на современных черепах, свидетельствуют о несомненно состоявшейся адаптации питекантропа к прямохождению. Общая массивность скелета питекантропа больше, чем у австралопитека. Многочисленные архаичные черты наблюдаются в строении черепа: сильноразвитый рельеф, покатый лобный отдел, массивные челюсти, выраженный прогнатизм лицевого отдела. Стенки черепа толстые, нижняя челюсть массивная и широкая, зубы крупные, при этом размеры клыка близки к современным. Сильноразвитый затылочный рельеф связан с развитием шейной мускулатуры, игравшей значительную роль в балансировке черепа при ходьбе. Приводимые в современной литературе оценки величины мозга питекантропов варьируют от 750 до 1350 см3, т. е. приблизительно соответствуют в минимуме нижнему порогу величин, приводимых для австралопитеков типа хабилисов. Ранее сопоставляемым видам приписывалось значительное отличие. Строение эндокранов свидетельствовало об усложнении структуры мозга: в большей степени у питекантропов развиты участки теменной области, нижнелобной и верхней задней части лобной области, что связывается с развитием специфических человеческих функций — труда и речи. На эндокранах синантропов были обнаружены новые очаги роста, связанные с оценкой положения тела, речи и тонких движений.
Синантроп несколько отличен по типу от питекантропа. Длина его тела составляла около 150 см (питекантропов — до 165-175 см), размеры черепа увеличены, но тип строения был тем же, исключение составлял ослабленный затылочный рельеф. Скелет синантропа менее массивен. Обращает на себя внимание грациальная нижняя челюсть. Объем мозга — более 1000 см3. Отличие синантропа и яванского питекантропа оценивается на уровне подвидового.
Характер пищевых остатков, как и строение нижних челюстей, указывает на изменение типа питания синантропов в сторону всеядности, что является прогрессивным признаком. У синантропов вероятен каннибализм. В вопросе об их способности добывать огонь археологи разошлись во мнениях.
Анализ костных остатков человека данной фазы антропогенеза позволяет реконструировать половозрастной состав коллективов синантропов: 3-6 самцов, 6-10 самок и 15-20 детей.
Сравнительная сложность культуры требует достаточно высокого уровня общения и взаимопонимания, следовательно, можно прогнозировать существование в это время примитивной речи. Биологическим основанием такого прогноза можно считать усиление костного рельефа в местах прикрепления мышц языка, начало формирования подбородка, грацилизацию нижних челюстей.
Фрагменты черепов древности, соизмеримой с ранними питекантропами о. Явы (примерно 1 млн лет), найдены в двух провинциях Китая — Ланьтянь, Куваньлинь. Интересно, что более древние китайские питекантропы отличаются от синантропов так же, как ранние питекантропы от поздних, а именно, большей массивностью костей и меньшей величиной мозга. К поздним прогрессивным питекантропам относится недавняя находка в Индии. Здесь вместе с позднеашельскими орудиями найден череп объемом 1300 см3.
Реальность существования стадии питекантропов в антропогенезе практически не оспаривается. Правда, предками последующих, более прогрессивных форм считаются поздние представители питекантропов. Широко обсуждался в науке вопрос о времени и месте появления первого питекантропа. Ранее его родиной считалась Азия, а время появления оценивалось примерно в 2 млн лет. Сейчас данный вопрос решается иначе. Родиной и австралопитеков, и питекантропов считается Африка. В 1984 г. в Кении (Нариокотоме) был открыт питекантроп древностью 1,6 млн лет (комплектный скелет подростка). Основными находками самых ранних питекантропов Африки считаются: Кооби-Фора (1,6 млн лет), южноафриканский Сварткранс (1,5 млн лет), Олдувай (1,2 млн лет). Африканские питекантропы побережья Средиземного моря (Тернифин) имеют древность 700 тыс. лет. Геологическую древность азиатских вариантов можно оценить в пределах 1,3-0,1 млн лет. Известны археологические свидетельства из памятников Ближнего Востока, находящихся ближе к Африке, чем к Азии, говорящие о том, что древность африканских питекантропов могла достигать 2 млн лет.
Синхронные формы ископаемого человека из Европы имеют меньший возраст и достаточно своеобразны. Их часто именуют «пренеандертальцами» или относят к Homo heidelbergensis, который в Африке, Европе и Азии был предковым для человека современного типа и неандертальцев Европы и Азии. Европейские формы имеют следующий возраст: Мауэр (500 тыс. лет), Араго (400 тыс. лет), Петралона (450 тыс. лет), Атапуэрка (300 тыс. лет). Переходный эволюционный характер имеют в Африке Брокен-Хилл (300 тыс. лет) и Бодо (600 тыс. лет).
На Кавказе наиболее древней считается находка в Грузии дманисского человека, древность которого оценивают в 1,6-1,8 млн лет. Анатомические особенности позволяют поставить его в один ряд с древнейшими гоминидами Африки и Азии! Питекантропы найдены и в других памятниках: в Узбекистане (Сель-Унгур), на Северном Кавказе (Кударо), Украине. Форма, промежуточная между питекантропами и неандертальцами, найдена в Азербайджане (Азых). Человек ашельского времени, видимо, жил на территории Армении (Ереван).
Ранние питекантропы отличаются от более поздних большей массивностью костей и меньшей величиной мозга. Подобное различие наблюдается в Азии и Европе.
В палеолите ашельская эпоха соотносится с физическим типом питекантропов и ранних неандертальцев. Ведущее орудие ашеля — ручное рубило (рис. I. 9). Оно демонстрирует высокий уровень в развитии технологии обработки камня. В пределах ашельской эпохи можно наблюдать увеличение тщательности отделки рубил: увеличивается количество сколов с поверхности орудия. Обработка поверхности становится более тонкой при смене каменных отбойников на более мягкие, сделанные из кости, рога или дерева. Размер ручного рубила достигал 35 см. Оно вырабатывалось из камня путем обработки сколами с двух сторон. Рубило имело заостренный конец, два продольных лезвия и необработанный противоположный край. Считается, что рубило имело разнообразные функции: оно служило ударным инструментом, использовалось для выкапывания кореньев, расчленения трупов животных, обработки дерева. В южных районах встречается топор (кливер), отличающийся поперечным лезвием, не подправленным ретушью, и симметрично обработанными краями.

Рис. I. 9. Ашельские ручные рубила
Типичное ашельское рубило не исчерпывает всего технологического разнообразия, характерного для того периода. Существовала отщеповая «клектонская» культура, а также отщеповая прогрессивная культура «леваллуа», которую отличает изготовление орудий из отщепов дисковидных камней-заготовок, поверхность заготовок предварительно обрабатывалась мелкими сколами. Кроме рубил, в ашельских памятниках встречаются мелкие орудия типа острий, скребков, ножей. Некоторые из них доживают до времени кроманьонцев. Встречаются в ашеле и олдувайские орудия. Известны редкие орудия из дерева. Предполагают, что питекантропы Азии могли обходиться орудиями из бамбука.
Большое значение в жизни ашельцев имела охота. Питекантропы не были только собирателями. Памятники ашеля интерпретируют как охотничьи стойбища, так как в их культурном слое встречаются кости крупных животных. Жизнь ашельских коллективов была сложной, люди занимались разными видами труда. Открыты разные типы стоянок: охотничьи лагеря, мастерские по добыче кремня, долговременные стоянки. Жилища ашельцы строили на открытых местах и в пещерах. В районе г. Ниццы открыто поселение из хижин.
Природное окружение ашельского человека определяло особенности материальной культуры. Типы орудий в разных памятниках встречаются в различных соотношениях. Охота на крупных животных требовала тесного сплочения коллектива людей. Стоянки разных типов свидетельствуют о существовании разделения труда. Остатки очагов говорят об эффективности использования питекантропами огня. В кенийской стоянке Чесованджа следы огня имеют древность 1,4 млн лет. Культура мустье неандертальского человека является развитием технологических достижений ангельской культуры питекантропов.
В результате афро-азитских маграций первых людей возникло два основных центра эволюции человека — западный и восточный. Разделенные огромными расстояниями популяции питекантропов могли прогрессировать длительное время в изоляции друг от друга. Существует мнение, что неандертальцы не были во всех регионах закономерной ступенью эволюции, в Африке и Европе таковой были питекантропы («пренеандертальцы»).
Неандертальцы (древние люди, палеоантропы)
В традиционной стадиальной модели антропогенеза промежуточную эволюционную ступень между Homo erectus и Homo sapiens представляли собою палеоантропы («древние люди»), которые в абсолютном летоисчислении обитали в период от 300 тыс. лет до примерно 30 тыс. лет в Европе, Азии и Африке. В непрофессиональной литературе они зачастую именуются «неандертальцами», по названию одной из первых находок 1848 г. в местности Неандерталь (Германия).
В общем, палеоантропы продолжают линию эволюции «Человека прямоходящего» (точнее — Homo heidelbergensis), но в современных схемах часто обозначаются как боковая ветвь гоминид. По общему уровню эволюционных достижений эти гоминиды наиболее близки к человеку современного вида. Поэтому они претерпели в классификациях гоминид изменения в своем статусе: палеоантропы в настоящее время рассматриваются как подвид «Человека разумного», т. е. как его ископаемый вариант (Homo sapiens neanderthalensls). Такой взгляд отражает новые знания о сложности биологии, интеллектуальной сферы и социальной организации неандертальцев. Антропологи, придающие большое значение биологическим отличиям неандертальца и современного человека, по-прежнему считают их особым видом.
Первые находки неандертальцев были сделаны в XIX в. в Западной Европе и не имели однозначной интерпретации.
Группы палеоантропов, разместившись в значительном диапазоне геологического времени, весьма многообразны по морфологическому облику. Антрополог В.П. Алексеев осуществил попытку классификации групп неандертальцев, сходных морфологически и хронологически, и выделил несколько группировок: европейскую, африканскую, типа Схул и переднеазиатскую. Известна большая часть находок палеоантропов из Европы. Часто неандертальцы заселяли приледниковые зоны.
На тех же основаниях (морфологическом и хронологическом) среди европейских форм указанного времени выделяют уровни: «самые ранние неандертальцы» — «пренеандертальцы», «ранние неандертальцы» и «поздние неандертальцы».
Антропологи предположили, что объективно существовали множественные переходы между последовательными стадиальными группами, поэтому в разных районах от нескольких вариантов питекантропов мог осуществиться эволюционный переход к палеоантропам. Представители вида Homo heidelbergensis могли быть предшественниками (Петралона, Сванскомб, Атапуэрка, Араго и др.).
В самую раннюю европейскую группу относят ископаемый череп из памятника Штейнгейм (200 тыс. лет), найденный на территории Германии в 1933 г., а также женский череп Сванскомб (200 тыс. лет), открытый в Англии в 1935 г. Данные находки относятся ко второму межледниковью по альпийской схеме. В аналогичных условиях, найдена ископаемая нижняя челюсть во Франции — памятник Монморен. Указанные формы отличаются небольшой величиной мозговой полости (Штейнгейм — 1150 см3, Сванскомб — 1250-1300 см3). Выделен комплекс признаков, сближающих самые ранние формы с современным человеком: сравнительно узкий и высокий череп, относительно выпуклый лоб, массивное надбровье, как у питекантропов, не разделенное на составляющие элементы, довольно округлый затылок, выпрямленный лицевой отдел, наличие зачаточного подбородка нижней челюсти. В строении зубов наблюдается явный архаизм: третий моляр по величине больше второго и первого (у человека размеры моляров уменьшаются от первого к третьему). Кости данной разновидности ископаемого человека сопровождены архаичными ашельскими орудиями.
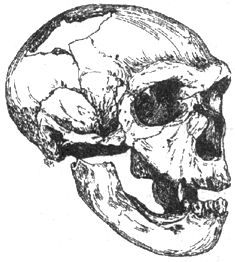
Рис. I. 10. Череп позднего европейского неандертальца (равняй вюрм)
К последнему межледниковью относятся многие известные науке неандертальцы. Более ранние из них жили примерно 150 тыс. лет назад. Представить их облик можно по находкам из европейских памятников Эрингсдорф и Саккопасторе. Их отличают вертикальный профиль лицевого отдела, округлый затылочный отдел, ослабленный надбровный рельеф, довольно выпуклый лоб, относительно небольшое число архаичных черт в строении зубов (третий моляр не самый крупный среди других). Объем мозга ранних неандертальцев оценивается в 1200-1400 см3.
Время существования поздних европейских неандертальцев совпадает с последним оледенением. Морфологический тип указанных форм хорошо виден на ископаемых костных остатках Шапелль (50 тыс. лет), Мустье (50 тыс. лет), Феррасси (50 тыс. лет), Неандерталь (50 тыс. лет), Энгис (70 тыс. лет), Чирчео (50 тыс. лет), Сан-Сезер (36 тыс. лет) (рис. I. 10).
Для данного варианта характерны сильное развитие надбровья, сжатый сверху вниз затылочный отдел («шиньонообразный»), широкое носовое отверстие, расширенная полость коренных зубов. Морфологи отмечают наличие затылочного валика, подбородочный выступ (редко и в зачаточной форме), большой объем мозговой полости: от 1350 до 1700 см3. По костям скелета туловища можно судить о том, что поздним неандертальцам было присуще сильное, массивное телосложение (длина тела — 155-165 см). Нижние конечности короче, чем у современного человека, бедренные кости изогнуты. Широкая лицевая часть черепа у неандертальцев сильно выступает вперед и скошена по бокам, скуловые кости обтекаемой формы. Суставы рук и ног велики. По пропорциям тела неандертальцы были похожи на современный тип эскимосов, что помогало им поддерживать температуру тела в условиях холодного климата.
Интересна попытка перенесения экологических знаний о современном человеке на палеоантропологические реконструкции. Так, ряд структурных особенностей «классических» неандертальцев Западной Европы объясняется следствием адаптации к условиям холодного климата.
Представляется, что самые ранние и последующие формы из Европы связаны генетическими связями. Европейские неандертальцы открыты на территории Франции, Италии, Югославии, Германии, Чехословакии, Венгрии, в Крыму и на Северном Кавказе.
Для решения вопроса о происхождении современного человека исключительно интересны находки палеоантропов за пределами Европы, главным образом, в Юго-Западной Азии и Африке. Отсутствие черт специализации в морфологии в большинстве случаев отличает их от европейских форм. Так, для них характерны более прямые и тонкие конечности, не столь мощные надглазничные валики, укороченные и менее массивные черепа.
Согласно одной из точек зрения, типичный неандертальский человек существовал лишь в пределах Европы и некоторых регионах Азии, куда он мог переселиться из Европы. Причем, начиная с рубежа 40 тыс. лет, неандертальцы сосуществовали с вполне сложившимися людьми современного анатомического типа; на Ближнем Востоке подобное сосуществование могло быть более длительным.
Исключительны по значению находки палеоантропов горы Кармел (Израиль). Они привлекли исследователей мозаикой сапиентных и неандерталоидных особенностей. Указанные находки можно трактовать как фактическое свидетельство метисации ранних неандертальцев и современного человека. Правда, надо заметить, что некоторые находки Схул в настоящее время рассматриваются как принадлежащие «архаичному Homo sapiens» . Назовем некоторые, наиболее известные находки.
Табун — ископаемый череп, открытый в пещере Табун, гора Кармел. Древность — 100 тыс. лет. Череп невысок, лоб покатый, есть надглазничные валики, но лицевая часть и затылочный отдел имеют современный характер. Изогнутые кости конечностей напоминают тип европейских неандертальцев.
Схул-V, древность — 90 тыс. лет (рис. I. 11). Череп сочетает большой объем мозговой полости и довольно высокий лоб с современным строением лицевого отдела и затылка.
Амуд, древность — 50 тыс. лет. Найден в пещере Амуд около Тивериадского оз. (Израиль). Имеет большой объем мозга: 1740 см3. Кости конечностей удлиненные.
Кафзех, древность — около 100 тые. лет. Открыт в Израиле. Сапиентность достаточно сильно выражена, поэтому считается состоявшимся сапиенсом.
На севере Ирака открыт шанидарский неандерталец, классический по типу, обладающий крупным мозговым отделом, исследователи обратили внимание на отсутствие сплошного надглазничного валика. Возраст — 70-80 тыс. лет.
На территории Узбекистана сделана находка неандертальца со следами погребального обряда. Череп принадлежал мальчику с неоформившимся надглазничным валиком. Лицевой отдел и конечности скелета, по мнению некоторых антропологов, имеют современный тип. Место находки — пещера Тещик-Таш, древность — 70 тыс. лет.
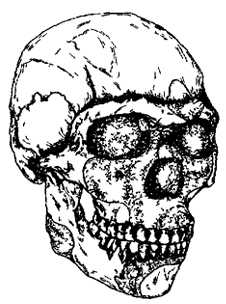
Рис. I. 11. Череп прогрессивного неандертальца (архаичного сапиенса) (90 тыс. лет)
В Крыму, в пещере Киик-Коба, были обнаружены костные остатки взрослого палеоантропа (тип близок к западноевропейским неандертальцам) и ребенка-неандертальца, очень малого возраста. Костные остатки нескольких неандертальских детей были открыты в Крыму и в районе г. Белогорска. Здесь же найден фрагмент черепа женщины-неандертальца с некоторыми современными особенностями, делающими ее похожей на схульские находки. Открыты неандертальские кости и зубы на территории Адыгеи и в Грузии.
Череп палеоантропа открыт в Азии — на территории Китая, в гроте Мала. Считается, что он не может быть отнесен ни к одному европейскому варианту неандертальцев. Важность данной находки заключается в том, что она доказывает замещение в азиатской части света одного стадиального типа другим. Другая точка зрения заключается в том, что в находках типа Мала, Чаньян, Ордос (Монголия) мы видим переходные формы от питекантропов к «ранним» сапиенсам. Причем, этот переход по некоторым формам можно датировать временем не менее 0,2 млн лет (урановый метод).
На о. Ява, в районе деревни Нган-донг, найдены своеобразные черепа, несущие следы каннибализма. Исследователи обратили внимание на их очень толстые стенки и мощный надглазничный валик. Такие признаки делают похожими нгандонгские черепа на тип питекантропов. Время существования открытых гоминид — верхний плейстоцен (около 0,1 млн лет), т. е. они синхронны поздним питекантропам. В науке существовало мнение, что это местный, своеобразный тип неандертальца, сформировавшийся в результате замедленного эволюционного процесса. С других позиций, «явантропы» из Нгандонга определяются как поздние питекантропы, генетически связанные с позднеплейстоценовыми сапиенсами Австралии.
До недавнего времени считалось, что неандертальцы существовали не только на севере, но и на юге Африки. В качестве примеров «южных» африканцев назывались гоминиды из Брокен-Хилл и Салданьи. В их морфологическом типе находили общие признаки неандертальцев и питекантропов. Объем мозга достигал у них около 1300 см3 (чуть меньше средней величины у неандертальцев). Высказывалось предположение, что человек из Брокен-Хилл является преемником олдувайского питекантропа из Восточной Африки. Некоторые антропологи считали, что существовала параллельная линия эволюции палеоантропов в Юго-Восточной Азии и на юге Африки. В настоящее время варианту Брокен-Хилл отведена роль ископаемой сапиентной формы.
Изменение таксономических взглядов на поздних гоминид привело к тому, что многие предшествующие современному человеку формы относят к архаичному Homo sapiens, часто понимая под этим термином «пронеандертальцев» (Сванскомб, Штейнгейм), далее — своеобразные африканские формы (Брокен-Хилл, Салданья), азиатские (Нгандонг), а также европейские варианты питекантропов.
Палеонтологические данные свидетельствуют о метисном происхождении классических европейских неандертальцев. Видимо, существовали две волны мигрантов из Африки и Азии примерно 300-250 тыс. лет назад, с последующим смешением.
Эволюционная судьба неандертальцев не ясна. Выбор гипотез достаточно широк: полная трансформация неандертальцев в сапиенсов; полное истребление неандертальцев сапиенсом внеевропейского происхождения; метисация обоих вариантов. Наибольшую поддержку имеет последняя точка зрения, согласно которой формирующийся человек современного типа мигрировал из Африки в Европу через Азию. В Азии он зафиксирован около 100 тыс. лет, а в Европу попал на рубеже 40 тыс. лет. Далее происходила ассимиляция неандертальского населения. Доказательством служат европейские находки гоминид неандертальского облика, современного типа и промежуточных форм. Ранние неандертальцы, проникая в Переднюю Азию, могли метисироваться с древним сапиенсом и там.
Представление о масштабах метисационных процессов дают ископаемые одонтологические материалы. На них зафиксирован вклад европейских неандертальцев в генофонд современного человека. Неандертальский вариант ископаемых гоминид сосуществовал с современным на протяжении десятков тысяч лет.
Сущность эволюционного перехода, имевшего место на границе верхнего палеолита, объяснена в гипотезе профессора Я.Я. Рогинского.
Автор обобщает данные о структуре эндокрана с клиническими наблюдениями современного человека и на этой основе выдвигает предположение, что социальное поведение палеоантропов и современного человека в значительной степени различается (контроль поведения, проявление агрессивности).
Мустьерская эпоха, совпадающая во времени с эпохой существования неандертальцев, относится к среднему палеолиту. В абсолютном исчислении данное время составляет от 40 до 200 тыс. лет. Мустьерские комплексы орудий неоднородны по соотношению орудий разного типа. Мустьерские памятники известны в трех частях света — Европе, Африке и Азии, там же открыты и костные остатки неандертальцев.
Технология обработки камня неандертальским человеком отличается относительно высоким уровнем техники раскалывания и вторичной обработки отщепов. Вершиной техники является прием подготовки поверхности камня-заготовки и обработки отделенных от него пластин.

Рис. 1.12. Мустьерские орудия среднего палеолита
Тщательная подправка поверхности заготовки влекла за собой тонкость пластин и совершенство получаемых из них орудий (рис. 1.12).
Для мустьерской культуры характерны дисковидные заготовки, от которых отщепы скалывались радиально: от краев к центру. Большая часть мустьерских орудий изготовлялась на отщепах путем вторичной обработки. Археологи насчитывают десятки типов орудий, но их разнообразие сводится, видимо, к трем видам: остроконечнику, скреблу, рубильцу. Острие представляло собою орудие с острием на конце, использовалось для разрезания мяса, кожи, обработки дерева, а также в качестве кинжала или наконечника копья. Скреблом служил отщеп, обработанный по краю ретушью. Данное орудие использовалось для скобления или резания при обработке туш, шкур или дерева. К скреблам добавлялись деревянные рукоятки. Зубчатые орудия применялись для обтачивания деревянных предметов, для резания или пиления. Встречаются в мустье проколки, резцы, скребки — орудия позднего палеолита. Средства труда представлены специальными отбойниками (куски камня или гальки удлиненной формы) и ретушерами (куски камня или кости для обработки края орудия путем нажима).
Современные этнографические исследования австралийских аборигенов помогают представить технологические процессы каменного века. Эксперименты археологов показали, что техника получения заготовок орудий в виде отщепов и пластин была сложной, требовала опыта, технических знаний, точной координации движений, большого внимания.
Опыт позволял древнему человеку сокращать количество времени, необходимого для изготовления орудий. Техника обработки кости в мустье развита слабо. Широко применялись орудия из дерева: дубины, копья, рогатины с закаленными на огне концами. Из дерева изготовлялись сосуды для воды и элементы жилищ.
Неандертальцы были искусными охотниками. На их стоянках обнаружены скопления костей крупных животных: мамонтов, пещерных медведей, бизонов, диких лошадей, антилоп, горных козлов. Сложные охотничьи действия были под силу согласованному коллективу неандертальцев. Мустьерцы использовали приемы облав или гона животных к обрывал и топям. Обнаружены сложносоставные орудия — наконечники копий с кремневыми осколками. Бола использовались как метательные орудия. Мустьерцы практиковали разрезание туш убитых животных и прожаривали мясо на огне. Они делали для себя простую одежду. Определенное значение имело собирательство. Обнаруженные зернотерки из камня позволяют предполагать, что существовала примитивная обработка зерна. Каннибализм у неандертальцев существовал, но не был широко распространен.
В мустьерское время изменился характер поселений. Чаще заселялись навесы, гроты и пещеры. Выделены типа неандертальских поселений: мастерские, охотничьи и базовые стоянки. Для защиты костров от ветра устраивались ветровые заслоны. В гротах делались вымостки из галек и кусков известняка.
Костные остатки неандертальцев могут быть найдены вместе с орудиями верхнего палеолита, как это было, например, при находке позднего палеоантропа во Франции (памятник Сен-Сезер).
В эпоху раннего вюрма на территории Евразии появились мустьерские погребения — первые достоверные следы захоронения умерших. Сегодня открыто около 60 таких памятников. Интересно, что «неандерталоидная» и «сапиентная» группы чаще погребали взрослых индивидов, а «неандертальское» население в одинаковой степени хоронило и взрослых, и детей. Факты захоронения умерших дают основание предполагать существование у мустьерцев дуалистического мировосприятия.
Человек современного типа, ископаемый и современный (неоантропы)
Ископаемые представители Homo sapiens sapiens широко представлены в известных археологических находках остатков гоминид. Максимальный геологический возраст полностью сформировавшихся в эволюции ископаемых неоантропов ранее оценивался примерно в 40 тыс. лет (находка в Индонезии). Сейчас считается, что сапиенс, найденный в Африке и Азии, имел гораздо большую древность (правда, речь идет о скелетах, имеющих архаичные признаки, выраженные в разной степени).
Костные остатки ископаемого человека данного подвида распространены широко: от Калимантана до оконечностей Европы.
Название «кроманьонцы» (так обозначают в литературе ископаемых неоантропов) обязано известному французскому памятнику верхнего палеолита Кро-Маньон. Строение черепа и скелета туловища ископаемых неоантропов в принципе не отличаются от человека современного типа, хотя его кости более массивны.
По данным анализа костного материала из позднепалеолитических погребений, средний возраст кроманьонцев составлял 30-50 лет. Такая же продолжительность жизни сохранилась до средних веков. Патология костей и зубов встречается реже, чем травмы (зубы кроманьонцев были здоровыми).
Признаки отличия черепов кроманьонцев от неандертальцев (рис. 1.13): менее выступающий вперед лицевой отдел, высокое выпуклое темя, высокий прямой лоб, округлый затылок, меньшие по размерам четырехугольные глазницы, меньше общие размеры черепа, сформирован подбородочный выступ черепа; надбровный валик отсутствует, челюсти развиты слабее, зубы имеют малую полость. Главное отличие кроманьонцев от неандертальцев — в строении эндокрана. Палеоневрологи считают, что в позднем антропогенезе развивались лобные отделы мозга, включающие центры контроля поведения. Были усложнены внутренние связи мозга, но общие размеры мозга несколько уменьшились. Кроманьонцы были выше ростом (169-177 см) и сложены менее грубо, чем неандертальцы.
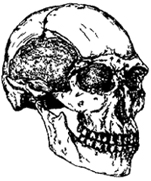
Рис. I. 13. Череп кроманьонца (Европа) (30 тыс. лет)
Отличия кроманьонских черепов от современных: меньше высота свода, больше продольные размеры, выражены надбровные дуги, больше ширина глазниц, шире лицевой отдел черепа и нижняя челюсть, больше толщина стенок черепа. Верхнепалеолитический человек достаточно долго сохранял признаки зубной системы, характерные для неандертальца. Признаки, отличающие череп и эндокран кроманьонцев от современного человека, по характеру часто являются «неандерталоидными».
Обращает на себя внимание тот факт, что ареал распространения кроманьонского человека огромен: вся ойкумена. С появлением кроманьонского человека, как считают многие специалисты, видовая эволюция человека завершается, а эволюция биологических качеств человеку в будущем представляется невозможной.
Наиболее комплектные находки скелетов кроманьонцев в Европе имеют древность, не превосходящую 40 тыс. лет. Например, французский неоантроп Кро-Маньон жил 30 тыс. лет назад, кроманьонец Сунгирь (район г. Владимира) имеет возраст 28 тыс. лет. Архаичный сапиенс Африки (с достаточно выраженными неандерталоидными особенностями) выглядит намного древнее: Омо в Эфиопии — 130 тыс. лет, Ривер Маус (Ю. Африка) — 120 тыс. лет, Бордер (Ю. Африка) — более 70 тыс. лет, кенийские находки сапиенса — 200-100 тыс. лет, Мумба (Танзания) — 130 тыс. лет и т. д. Предполагается, что древность африканского сапиенса может быть еще больше. Азиатские находки сапиенса имеют следующий возраст: Дали (КНР) — 200 тыс. лет, Цзиньнбшань (КНР) — 200 тыс. лет, Кафзех (Израиль) — более 90 тыс. лет, Схул V (Израиль) — 90 тыс. лет, Ниа (Калимантан) — 40 тыс. лет. Австралийские находки имеют древность около 10 тыс. лет.
Ранее предполагалось, что человек современного типа возник в Европе примерно 40 тыс. лет назад. Сегодня большее число антропологов и археологов помещают прародину сапиенса в Африку, а древность последнего сильно увеличивают, ориентируясь на приведенные выше находки. В соответствии с гипотезой немецкого антрополога Г. Бройера, Homo sapiens sapiens появился южнее Сахары примерно 150 тыс. лет назад, затем мигрировал в Переднюю Азию (на уровне 100 тыс. лет), а на рубеже 35-40 тыс. лет стал заселять Европу и Азию, метисируясь с местными неандертальцами. Современные биомолекулярные данные также позволяют предположить, что предки современного человечества — выходцы из Африки.
В соответствии с современными эволюционными воззрениями, наиболее правдоподобной представляется модель «сетчатой эволюции» гоминид, в которой важное место отведено обмену генами между разными подвидами и видами древнего человека. Поэтому очень ранние находки сапиенса в Африке и Европе трактуют как свидетельство метисации между видами сапиенса и питекантропами. В процессе становления сапиентного типа между первичными центрами эволюции рода Homo (западным и восточным) происходил постоянный обмен генами.
Около 40 тыс. лет назад началось стремительное расселение неоантропа. Причины данного феномена заключены в генетике человека и развитии его культуры.
Ученым, занимающимся изучением кроманьонского человека, приходится сталкиваться с большим разнообразием его типов. Единого мнения о времени сложения современных рас нет. Согласно одной точке зрения, черты современных рас есть в верхнем палеолите. Эту точку зрения иллюстрируют примерами географического распространения двух признаков — выступания носа и степени горизонтальной профилировки лицевого отдела. Согласно другой точке зрения, расы оформляются поздно, а население верхнего палеолита отличалось большим полиморфизмом. Так, для Европы иногда выделяют около 8 типов рас верхнего палеолита. Два из них выглядят так: а) долихокранный, большеголовый вариант кроманьонца с умеренной шириной лица и узким носом; б) брахикранный (короткоголовый), с меньшими размерами черепа, очень широким лицом и широким носом. Можно предположить, что существовало три этапа формирования рас: 1) средний и нижний палеолит — образование некоторых расовых черт; 2) верхний палеолит — начало формирования расовых комплексов; 3) послепалеолитическое время — сложение рас.
Культуры верхнего (позднего) палеолита связаны с появлением человека современного типа (неоантропа). В Европе последний период палеолита (древнего каменного века) оценивается временем 35-10 тыс. лет до наших дней и совпадает со временем последнего плейстоценового оледенения (этот факт является предметом обсуждения в связи с проблемой роли окружающей среды в развитии человечества) (рис. I. 14).
На первый взгляд, в обсуждаемую эпоху палеолита отсутствовали кардинальные отличия материальной культуры от предшествующих эпох: те же каменные инструменты и охотничьи орудия. На самом деле, кроманьонцы изготавливали более сложный набор орудий: ножи (иногда кинжалы), наконечники копий, резцы долота, костяные орудия типа шильев, игл, гарпунов и т. д. Костяные орудия составляли около половины всего инвентаря, они были прочными и более долговечными, чем каменные. Каменные орудия применялись для выделки орудий из кости, дерева, слоновой кости — так были усложнены технологические цепи в действиях древнего человека.

Рис. I. 14. Орудия верхнего палеолита
Возникли совершенно новые типы орудий, такие, как иглы с ушками, рыболовные крючки, гарпуны, копьеметалки. Они существенно усилили власть человека над природой.
Главное отличие верхнего палеолита заключалось в совершенствовании обработки камня. В мустьерское время существовало несколько способов обработки камня-заготовки (нуклеуса). Лаваллузская техника тщательной первоначальной обработки поверхности заготовки — старт для техники верхнего палеолита. Кроманьонцы использовали заготовки, пригодные для скалывания серий пластин (призматические нуклеусы). Таким образом, в эпоху верхнего палеолита совершенствовалась техника скалывания, в результате чего получались микропластины высокого качества, пригодные для использования в составных орудиях.
Археологи провели эксперименты по реконструкции способа отделения пластин от нуклеуса, как это делали кроманьонцы. Отобранный и специально обработанный нуклеус зажимался между колен, игравших роль амортизатора. Отделение пластин производилось при помощи отбойника из камня и посредника из кости. Кроме того, кремневые пластины отделялись путем нажима на край ядрища костяным или каменным отжимником.
Метод ножевидных пластин много экономичнее метода отщепов. От одной заготовки умелый мастер мог отделить за короткое время более 50 пластин (длина до 25-30 см, а толщина — несколько миллиметров). Рабочий край ножевидной пластины много больше, чем у отщепа. Для позднего палеолита известно более 100 типов орудий. Высказано предположение, что разные мастерские кроманьонцев могли отличаться своеобразием технической «моды».
В верхнем палеолите охота была еще более совершенной, чем в мустьерское время. Это играло огромную роль в увеличении пищевых ресурсов, и, в связи с этим, населения.
Совершенным новшеством явились копьеметалки, которые давали руке кроманьонца выигрыш в силе, удваивая расстояние, на которое можно было метнуть копье (до 137 м, с оптимальным для поражения расстоянием до 28 м). Гарпуны позволяли эффективно добывать рыбу. Кроманьонец изобрел силки для птиц, ловушки для зверей.
Совершенная охота осуществлялась на крупного зверя: северный олень и козероги преследовались во время их сезонных миграций на новые пастбища и обратно. Приемы охоты с использованием знания местности — загонная охота — позволяли убивать зверей тысячами. Так впервые сформировался бесперебойный источник высокопитательной пищи. Человек получил возможность проживания в труднодоступных районах.
В строительстве жилищ кроманьонцы использовали достижения мустьерцев и усовершенствовали их. Это позволило им выжить в условиях последнего холодного тысячелетия плейстоцена.
Европейские кроманьонцы, заселяя пещеры, использовали хорошее знание местности. Многие пещеры имели выход на юг, поэтому хорошо обогревались солнцем и были защищены от холодных северных ветров. Пещеры выбирались неподалеку от источников воды, с хорошим обзором пастбищ, где паслись стада копытных. Пещеры могли использоваться круглый год или для сезонного пребывания.
Кроманьонцы строили жилища также в речных долинах. Их складывали из камня или вырывали в земле, стены и крыша изготовлялись из шкур, а подпорки и низ могли обкладываться тяжелыми костями и бивнями. Верхнепалеолитическое строение в местонахождении Костенки (Русская равнина), длиной 27 м, отмечено рядом очагов по центру, что свидетельствует о том, что здесь зимовали несколько семей.
Кочующие охотники строили легкие шалаши. Суровые климатические условия кроманьонцам помогала переносить теплая одежда. Изображения людей на костяных поделках позволяют представить, что они носили тесно прилегающие штаны, сохраняющие тепло, парки с капюшонами, обувь и рукавицы. Швы одежды хорошо прошивались.
Высокое интеллектуальное развитие и психологическая сложность кроманьонцев доказаны существованием многочисленных памятников первобытного искусства, которое известно для периода 35-10 тыс. лет в Европе. Имеются в виду скульптуры малых форм и настенная живопись в пещерах. Гравюры животных и людей были сделаны на камнях, костях и оленьих рогах. Скульптуры и барельефы изготовлялись из глины и камня, а рисунки получены кроманьонцами с помощью охры, марганца и древесного угля. Предназначение первобытного искусства не ясно. Предполагают, что оно носило ритуальный характер.
Обильную информацию о жизни кроманьонцев дают исследования погребений. Выяснено, например, что продолжительность жизни кроманьонского человека в сравнении с неандертальцами возросла.
Реконструированы некоторые ритуалы кроманьонцев. Так, обычай посыпать скелет умершего красной охрой свидетельствует, видимо, о вере в загробную жизнь. Погребения с богатыми украшениями позволяют предположить появление в среде охотников-собирателей богатых людей.
Прекрасный пример кроманьонского погребения дает памятник Сунгирь под г. Владимиром. Возраст погребения — около 24-26 тыс. лет. Здесь покоится старик («Вождь») в меховых одеждах, богато украшенных бусами. Интересно второе погребение — парное детское. Скелеты детей сопровождались копьями из бивней мамонта и были украшены кольцами и браслетами из слоновой кости; одежды также украшены бусинами.
Современный человек и эволюция
Со времени завершения формирования вида Homo sapiens (с середины верхнего палеолита) он сохраняет в своем биологическом статусе стабильность. Эволюционная завершенность человека относительна и не означает полного прекращения изменений его биологических свойств. Изучены разнообразные изменения анатомического типа человека современного типа. Примерами служат уменьшение массивности скелета, размеров зубов, изменение малых пальцев стопы и т. д. Предполагается, что данные явления обусловлены случайными мутациями. Некоторые антропологи на основании анатомических наблюдений прогнозируют появление Homo futurus — «Человека будущего», с крупной головой, уменьшенным лицом и зубами, с меньшим количеством пальцев. Но указанные анатомические «утраты» не характеризуют всех популяций человека. Альтернативный взгляд заключается в том, что биологическая организация современного человека дает возможность безграничной социальной эволюции, поэтому вряд ли в будущем он изменится как вид.
К содержанию учебника «Антропология» | К следующей главе